Which prompts the question: if genes contributing to group differences in IQ is so awful and horrible and terrible, why isn’t it similarly awful that genes contribute to individual differences in IQ? (This question is directed more at people like Paige Harden than at Grossman, who’s obviously not an expert in this area and doesn’t claim to be.)
…
The idea that there’s a sharp distinction between individual and group differences doesn’t make any sense. Either it’s bad to claim that some humans are “just inherently dumber” than other humans, or it isn’t.
In fact, this general take goes back a long time, and it’s worth expanding upon. Pretty much everything is this field has already been said by Arthur Jensen somewhere in his 400+ papers and 5+ books. This particular idea is no different. Here’s Jensen in 1998:
The relationship of the g factor to a number of biological variables and its relationship to the size of the white-black differences on various cognitive tests (i.e., Spearman’s hypothesis) suggests that the average white-black difference in g has a biological component. Human races are viewed not as discrete, or Platonic, categories, but rather as breeding populations that, as a result of natural selection, have come to differ statistically in the relative frequencies of many polymorphic genes. The “genetic distances” between various populations form a continuous variable that can be measured in terms of differences in gene frequencies. Racial populations differ in many genetic characteristics, some of which, such as brain size, have behavioral and psychometric correlates, particularly g. What I term the default hypothesis states that the causes of the phenotypic differences between contemporary populations of recent African and European descent arise from the same genetic and environmental factors, and in approximately the same magnitudes, that account for individual differences within each population. Thus genetic and environmental variances between groups and within groups are viewed as essentially the same for both populations. The default hypothesis is able to account for the present evidence on the mean white-black difference in g. There is no need to invoke any ad hoc hypothesis, or a Factor X, that is unique to either the black or the white population. The environmental component of the average g difference between groups is primarily attributable to a host of microenvironmental factors that have biological effects. They result from non genetic variation in prenatal, perinatal, and neonatal conditions and specific nutritional factors.
This is in fact his summary of the 12th chapter in his book where he makes the case for the genetic factors in explaining American race gaps, chiefly the Black-White one (the best studied). And yes, you should read the book. Even though it’s 25 years old, it is the best book on intelligence ever written.

The unification of individual and group differences is a reductionist approach to the problem. Instead of having one theory to explain individual variation in intelligence, and another unrelated theory to explain between group differences, it makes more sense to have a single theory of both. Jensen wisely proposes this should be considered the default on epistemic grounds, i.e., the prior in Bayesian terms. Take an example. Suppose you know that variation in height between plants is mainly caused by genetics and some soil differences. Next up you notice that the plants can be usefully grouped by the color and shape of their flowers. You measure their heights and it turns out that the oval yellow flowered plants are taller than the round orange flowered plants. Why is that, you ask. If you wanted to research this question, it would be very odd, indeed epistemically unwise, to begin with the hypothesis that this difference between the orange and yellow flowered plants are due to only soil differences. This isn’t technically impossible, but it is not a good starting point.
Jensen expands on his default hypothesis:
Consider the following items of evidence: the many biological correlates of g; the fact that among all of the psychometric factors in the domain of cognitive abilities the g factor accounts for the largest part of the mean difference between blacks and whites; the evolutionary history of Homo sapiens and the quantitative differentiation of human populations in allele frequencies for many character istics, including brain size, largely through adaptive selection for fitness in highly varied climates and habitats; the brain evolved more rapidly than any other organ; half of humans’ polymorphic genes affect brain development; the primary evolutionary differentiation and largest genetic distance between human populations is that between the African populations and all others; the intrinsic positive correlation between brain size and measures of g\ the positive mean white-black difference in brain size; the positive correlation between the variable heritability of individual differences in various measures of cognitive abilities and the variable magnitudes of their g loadings. All these phenomena, when viewed together, provide the basis for what I shall call the default hypothesis concerning the nature of population or racial differences in g.
Although we are concerned here with variation between populations, it is also important to keep in mind that, from an evolutionary perspective, it is most unlikely that there are intraspecies differences in the basic structural design and operating principles of the brain. The main structural and functional units of the brain found in any one normal human being should be validly generalizable to all other normal humans. That is to say, the processes by which the brain perceives, learns, reasons, remembers, and the like are the same for everyone, as are the essential structures and functions of every organ system in the entire body. Individual differences and population differences in normal brain processes exist at a different level, superimposed, as it were, over and above the brain’s common structures and operating principles.
The default hypothesis states that human individual differences and population differences in heritable behavioral capacities, as products of the evolutionary process in the distant past, are essentially composed of the same stuff, so to speak, controlled by differences in allele frequencies, and that differences in allele frequencies between populations exist for all heritable characteristics, physical or behavioral, in which we find individual differences within populations.
With respect to the brain and its heritable behavioral correlates, the default hypothesis holds that individual differences and population differences do not result from differences in the brain’s basic structural operating mechanisms per se, but result entirely from other aspects of cerebral physiology that modify the sensitivity, efficiency, and effectiveness of the basic information processes that mediate the individual’s responses to certain aspects of the environment.1271 A crude analogy would be differences in the operating efficiency (e.g., miles per gallon, horsepower, maximum speed) of different makes of automobiles, all powered by internal combustion engines (hence the same operating mechanisms) but differing in, say, the number of cylinders, their cubic capacity, and the octane rating of the gasoline they are using. Electric motor cars and steam-engine cars (analogous to different species or genera) would have such distinctively different operating mechanisms that their differences in performance would call for quite different explanations.
In other words, we don’t need to posit any mysterious extra causes (“I had no need of that hypothesis”). What Jensen didn’t realize in 1998 is that this default hypothesis is testable, indeed, provable with structural equation modeling. Just 5 years later, in a relatively unknown paper:
- Lubke, G. H., Dolan, C. V., Kelderman, H., & Mellenbergh, G. J. (2003). On the relationship between sources of within-and between-group differences and measurement invariance in the common factor model. Intelligence, 31(6), 543-566.
Investigating sources of within- and between-group differences and measurement invariance (MI) across groups is fundamental to any meaningful group comparison based on observed test scores. It is shown that by placing certain restrictions on the multigroup confirmatory factor model, it is possible to investigate the hypothesis that within- and between-group differences are due to the same factors. Moreover, the modeling approach clarifies that absence of measurement bias implies common sources of within- and between-group variation. It is shown how the influence of background variables can be incorporated in the model. The advantages of the modeling approach as compared with other commonly used methods for group comparisons is discussed and illustrated by means of an analysis of empirical data.
The authors spell it out:
The central issue of the present paper concerns the relation between MI on the one hand and within- and between-group differences on the other hand. Specifically, the definition of MI across groups implies that between-group differences cannot be due to factors with a different conceptual interpretation than the factors that account for the within-group differences. Although the importance of MI has been acknowledged (Byrne, Shavelson, & Muthén, 1989; Dolan, 2000; Lubke, Dolan, & Kelderman, 2001; Marsh, 1994; McArdle, 1998; Oort, 1998), this implication is not well recognized. Hence, if in practice the hypothesis of MI is not rejected, one can conclude with some confidence that within- and between-group differences are attributable to the same factors.
(Jensen died in 2012, but never cited this 2003 paper, so it appears he never learned about it, sad!)
The kind of measurement invariance they are talking about is called strict measurement invariance, which is where the intercepts and residuals are held constant. When this model is tenable, it means that there are no outside causes of variation in tests between one group and another. In simpler terms, there cannot be a cause of variance that makes one group better at, say, vocabulary, without also making the other group better. This finding is subtle but it rules out the whole class of possible options called the X factor. This is a hypothetical cause of intelligence test variation that only affects one group, either elevating it or depressing it. You can see how this idea is attractive to egalitarians because it seemingly allows them to posit a cultural racism factor that depresses Black people’s scores. But in fact it doesn’t work as James Flynn explained in 1987:
The two steps of Jensen’s case lend one another a logical force that each lacks in isolation. Usually he begins with his high h2 estimates because after all, an estimate of .80 means environmental factors play a limited role in explaining IQ differences within the black community. Assume that blacks in general suffer from an environmental handicap so potent as to reduce their mean IQ by fifteen points. Now if that environmental handicap affects some blacks more than others, its very potency would guarantee that it would account for much of the IQ differences within the black community. Therefore, we are forced to conclude that it affects every black to almost the same degree. But how probable is this? For example, take racism as the most likely factor that depresses black IQ below white. Racism may well handicap blacks through low motivation, unfavourable self-image, emasculation of the male, the welfare mother home. But certainly some blacks have drive, self-confidence, a stable home, so how can anyone argue that such factors affect blacks to almost the same degree? Unable to find a factor he can specify without embarrassment, the environmentalist is driven to assume a mysterious factor X, a sort of blindfold with no name, that must handicap every black and leave every white unscathed.
In other words, this is a logically possible — it’s not literally self-contradictory — but empirically hopeless idea. There’s no cause known in social science that doesn’t vary between people. Indeed, many egalitarians have spent many years trying to prove skin color discrimination. They do this by examining the outcomes of people of mixed ancestry who vary in skin color, and then show that those who are darker are worse off. Entire books are written about this “pigmentocracy” discrimination idea. Obviously, since skin color varies within groups, this cannot be an X factor. (The hereditarian reply is that skin color is a proxy for ancestry and thus intelligence and that’s why it predicts stuff.) But as Lubke et al later showed, even unknown X factors can be ruled out by finding strict measurement invariance. That’s why we have been spending quite a bit of time looking at large datasets to see if strict invariance holds, and it does:
- Hu, M., Lasker, J., Kirkegaard, E. O., & Fuerst, J. G. (2019). Filling in the gaps: The association between intelligence and both color and parent-reported ancestry in the National Longitudinal Survey of Youth 1997. Psych, 1(1), 240-261.
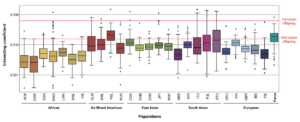
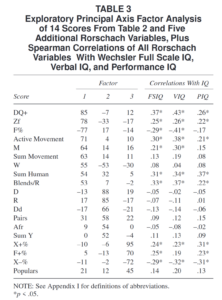
Little research has dealt with intragroup ancestry-related differences in intelligence in Black and White Americans. To help fill this gap, we examined the association between intelligence and both color and parent-reported ancestry using the NLSY97. We used a nationally-representative sample, a multidimensional measure of cognitive ability, and a sibling design. We found that African ancestry was negatively correlated with general mental ability scores among Whites (r = −0.038, N = 3603; corrected for attenuation, rc = −0.245). In contrast, the correlation between ability and parent-reported European ancestry was positive among Blacks (r = 0.137, N = 1788; rc = 0.344). Among Blacks, the correlation with darker skin color, an index of African ancestry, was negative (r = −0.112, N = 1455). These results remained with conspicuous controls. Among Blacks, both color and parent-reported European ancestry had independent effects on general cognitive ability (color: β = −0.104; ancestry: β = 0.118; N = 1445). These associations were more pronounced on g-loaded subtests, indicating a Jensen Effect for both color and ancestry (rs = 0.679 to 0.850). When we decomposed the color results for the African ancestry sample between and within families, we found an association between families, between singletons (β = −0.153; N = 814), and between full sibling pairs (β = −0.176; N = 225). However, we found no association between full siblings (β = 0.027; N = 225). Differential regression to the mean results indicated that the factors causing the mean group difference acted across the cognitive spectrum, with high-scoring African Americans no less affected than low-scoring ones. We tested for measurement invariance and found that strict factorial invariance was tenable. We then found that the weak version of Spearman’s hypothesis was tenable while the strong and contra versions were not. The results imply that the observed cognitive differences are primarily due to differences in g and that the Black-White mean difference is attributable to the same factors that cause differences within both groups. Further examination revealed comparable intraclass correlations and absolute differences for Black and White full siblings. This implied that the non-shared environmental variance components were similar in magnitude for both Blacks and Whites.
This finding was replicated in the PNC dataset, and in the NLSY79 and VES.
With the X factor defeated, the egalitarian must fall back to positing a cause that varies between people, but which affects either Black or Whites more. There are many such plausible ideas, but none of them are in fact sufficient. The problem is that the high heritability within groups demands a really powerful non-genetic cause that must show extreme variation between groups. No such cause exists however. My recent post went into detail about this.
The Newtonian analogy
Finally, it is worth pointing out that there is a historical analogy to this situation, namely Newton’s unification of the causes of the heavenly realm and the earthly realm:
Kepler had proposed three Laws of Planetary motion based on the systematics that he found in Brahe’s data. These Laws were supposed to apply only to the motions of the planets; they said nothing about any other motion in the Universe. Further, they were purely empirical: they worked, but no one knew a fundamental reason WHY they should work.
Newton changed all of that. First, he demonstrated that the motion of objects on the Earth could be described by three new Laws of motion, and then he went on to show that Kepler’s three Laws of Planetary Motion were but special cases of Newton’s three Laws if a force of a particular kind (what we now know to be the gravitational force) were postulated to exist between all objects in the Universe having mass. In fact, Newton went even further: he showed that Kepler’s Laws of planetary motion were only approximately correct, and supplied the quantitative corrections that with careful observations proved to be valid.
Before Newton, the causes of these realms were considered separate, mostly based on old Greek speculation. But they were in fact one and the same: no planetary X factors were needed to explain their movements.
Conclusion
We don’t need to posit any mysterious causes that only act on some groups of people to explain their intelligence levels. All human variation in intelligence has approximately the same causes. It happens that the positive alleles for intelligence are more common in some groups of people — people who earn a lot of money, people who read a lot of books, and in some ancestries — than they are in other groups of people. Indeed, it would be a miracle if they just happened to be exactly identical in their averages. There is no cosmic egalitarian God. The egalitarian science on this topic is a “environment of the gaps” where increasingly implausible and difficult to disprove causes are proposed to avoid the obvious yet undesirable conclusion about the role of genetics.
Extra: Historical precedent
Back in 1973 Jensen was already arguing about the unity of group and individual differences, he just didn’t label it the default hypothesis yet. Jensen was trying to reconcile findings from one of those foster care children studies showing large IQ gains with high heritability (Skodak and Skeels 1945, 1949). And there was a paper from the 1990s redoing this math and arguing the same point. Interestingly, that person was… Eric Turkheimer! So Eric knows that group differences and individual differences cannot be properly separated. Lately, he seems to have taken the even more extreme line that we shouldn’t talk about the causality of genetics insofar as we are analyzing polygenic scores. I assume that’s because he wants to block the link to causality and polygenic scores for group differences.
- Jensen, A. R. (1973a). Educability and group differences. New York: Harper & Row
- Turkheimer, E. (1991). Individual and group differences in adoption studies of IQ. Psychological Bulletin, 110(3), 392.
Extra: the super X factor
There is one way one can still have an X factor that isn’t ruled out by strict measurement invariance. However, this X factor must work exactly the same as the g factor and other non-g abilities in the model, i.e. have the same factor loadings, OR, it must work through the g factor, i.e. causing intelligence itself. So it would have to be something that doesn’t vary at all between Blacks (or Whites), and perfectly mimics the effects of real intelligence, but isn’t, or directly influence intelligence. It’s an even more fantastically improbable idea. For more on the X factor and this variant, see Dalliard’s 2014 post.